1. Introduction
The liver is particularly vulnerable to oxidative stress due to its central role in xenobiotic metabolism, detoxification, and energy homeostasis. As the principal organ responsible for biotransformation of drugs, environmental toxins, and metabolic by-products, hepatocytes are continuously exposed to reactive oxygen species (ROS) generated both during normal metabolic processes and through cytochrome P450-mediated xenobiotic metabolism, making them susceptible to oxidative injury. Excessive ROS production during detoxification disrupts redox balance, damages lipids, proteins, and DNA, and contributes to the pathogenesis of various liver diseases. This susceptibility is compounded by the liver’s high metabolic activity and its ongoing requirement to maintain redox homeostasis, positioning oxidative stress as a key factor in hepatic dysfunction and disease progression
| [10] | Conde de la Rosa, L., Goicoechea, L., Torres, S., Garcia-Ruiz, C., & Fernandez-Checa, J. C. (2022). Role of oxidative stress in liver disorders. Livers, 2(4), 283-314.
https://doi.org/10.3390/livers2040023 |
[10]
.
Oxidative stress is widely recognized as a fundamental biochemical mechanism underlying the initiation and progression of liver diseases and numerous chronic conditions, including metabolic syndrome, inflammation, cancer, and neurodegenerative disorders
| [28] | Sies, H., Berndt, C., & Jones, D. P. (2017). Oxidative stress. Annual Review of Biochemistry, 86, 715-748.
https://doi.org/10.1146/annurev-biochem-061516-045037 |
| [30] | Valko, M., Leibfritz, D., Moncol, J., Cronin, M. T., Mazur, M., & Telser, J. (2007). Free radicals and antioxidants in normal physiological functions and human disease. International Journal of Biochemistry & Cell Biology, 39(1), 44-84.
https://doi.org/10.1016/j.biocel.2006.07.001 |
[28, 30]
. It arises from an imbalance between excessive production of reactive oxygen species (ROS) and the inability of endogenous antioxidant defense systems to neutralize these reactive intermediates
. Under physiological conditions, ROS play essential roles in cellular signaling and metabolic regulation; however, their uncontrolled accumulation results in oxidative damage to biomolecules such as lipids, proteins, and DNA, ultimately impairing cellular integrity and function
| [4] | Betteridge D. J. (2000). What is oxidative stress?. Metabolism: Clinical and Experimental, 49(2 Suppl 1), 3-8.
https://doi.org/10.1016/s0026-0495(00)80077-3 |
| [14] | Halliwell, B., & Gutteridge, J. M. C. (2015). Free radicals in biology and medicine (5th ed.). Oxford University Press. |
[4, 14]
.
In vitro oxidative stress models using human hepatocellular carcinoma (HepG2) cells have become a widely accepted experimental platform for investigating hepatic redox imbalance and evaluating the antioxidant potential of bioactive compounds. Hydrogen peroxide (H
2O
2), a stable ROS precursor, is commonly employed to induce oxidative injury in HepG2 cells, mimicking pathological oxidative conditions observed in liver dysfunction
| [15] | Jaeschke, H. (2011). Reactive oxygen and mechanisms of inflammatory liver injury: Present concepts. Journal of Gastroenterology and Hepatology, 26(s1), 173-179.
https://doi.org/10.1111/j.1440-1746.2010.06592.x |
| [21] | Li, S., Tan, H. Y., Wang, N., Zhang, Z. J., Lao, L., Wong, C. W., & Feng, Y. (2015). The role of oxidative stress and antioxidants in liver diseases. International Journal of Molecular Sciences, 16(11), 26087-26124.
https://doi.org/10.3390/ijms161125942 |
[15, 21]
. In recent years, increasing attention has been directed toward plant-derived natural antioxidants as safer alternatives to synthetic antioxidants, which may exhibit adverse toxicological effects when used long-term. Medicinal plants and dietary vegetables are rich sources of phenolics, flavonoids, and other secondary metabolites capable of scavenging free radicals, chelating metal ions, and enhancing endogenous antioxidant enzyme activities. These phytochemicals contribute significantly to cellular defense mechanisms by acting as free radical scavengers, while also modulating key cellular signaling pathways
| [7] | Carlsen, M. H., Halvorsen, B. L., Holte, K., Bøhn, S. K., Dragland, S., Sampson, L., Willey, C., Senoo, H., Umezono, Y., Sanada, C., Barikmo, I., Berhe, N., Willett, W. C., Phillips, K. M., Jacobs, D. R., & Blomhoff, R. (2010). The total antioxidant content of more than 3100 foods, beverages, spices, herbs and supplements used worldwide. Nutrition Journal, 9(1), 3.
https://doi.org/10.1186/1475-2891-9-3 |
| [18] | Kensler, T. W., Wakabayashi, N., & Biswal, S. (2007). Cell survival responses to environmental stresses via the Keap1-Nrf2-ARE pathway. Annual Review of Pharmacology and Toxicology, 47, 89-116.
https://doi.org/10.1146/annurev.pharmtox.46.120604.141046 |
| [31] | Zhang, Y. J., Gan, R. Y., Li, S., Zhou, Y., Li, A. N., Xu, D. P., & Li, H. B. (2015). Antioxidant phytochemicals for the prevention and treatment of chronic diseases. Molecules, 20(12), 21138-21156. https://doi.org/10.3390/molecules201219753 |
[7, 18, 31]
.
Beta vulgaris (beetroot) is a widely consumed vegetable known for its high nutritional and pharmacological value. It contains abundant bioactive compounds, including betalains, polyphenols, flavonoids, carotenoids, and ascorbic acid, which have been reported to exhibit strong antioxidant, anti-inflammatory, hepatoprotective, and cytoprotective activities. Betalains, in particular, are potent radical scavengers and have been associated with reduced lipid peroxidation and improved cellular redox balance
| [9] | Clifford, T., Howatson, G., West, D. J., & Stevenson, E. J. (2015). The potential benefits of red beetroot supplementation in health and disease. Nutrients, 7(4), 2801-2822.
https://doi.org/10.3390/nu7042801 |
| [20] | Kujawska, M., Ignatowicz, E., Murias, M., Ewertowska, M., Mikolajczyk, K., & Jodynis-Liebert, J. (2009). Protective effect of red beetroot against carbon tetrachloride-induced oxidative stress in rats. Journal of Agricultural and Food Chemistry, 57(6), 2570-2575. https://doi.org/10.1021/jf803702d |
[9, 20]
. This study aims to evaluate the
in vitro antioxidant and protective effects of Beta vulgaris ethanol extract against H
2O
2-induced oxidative stress in HepG2 cells by assessing intracellular ROS generation, lipid peroxidation, and antioxidant enzyme activities.
2. Materials and Methods
Collection and Identification of Plant Sample Materials
Fresh Beta vulgaris (beetroot) roots were purchased from the Fruit Garden Market in Port Harcourt, Rivers State, Nigeria. The plant materials were subsequently authenticated at the Department of Plant Science and Biotechnology, Rivers State University, Nigeria.
2.1. Chemicals and Reagents
Analytical-grade ethanol, hydrogen peroxide (H2O2), 3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide (MTT), 2′, 7′-dichlorofluorescin diacetate (DCFH-DA), thiobarbituric acid (TBA), Ellman’s reagent (DTNB), and other biochemical assay reagents were purchased from standard commercial suppliers (Sigma-Aldrich, USA). Dulbecco’s Modified Eagle Medium (DMEM), fetal bovine serum (FBS), trypsin-EDTA, and penicillin–streptomycin were from Gibco (Thermo Fisher Scientific, USA).
2.2. Extraction and Preparation of Plant Materials
The fleshy bulbous roots of Beta vulgaris were thoroughly washed with distilled water to remove soil particles and other surface impurities. About 200g of cleaned plant material was then homogenized into fine slurry using a laboratory blender. The homogenate was macerated in 95% ethanol at a ratio of 1:5 (w/v) for 72 hours at room temperature with intermittent shaking to enhance the extraction of phytochemical constituents. After maceration, the mixture was first filtered through muslin cloth to remove coarse residues and subsequently through Whatman No. 1 filter paper to obtain a clear filtrate. The extraction process was repeated under identical conditions to maximize yield. The filtrates obtained from both extraction cycles were pooled together and concentrated to dryness using a water bath maintained at 45°C. The resulting crude ethanol extract was dried completely and stored at 4°C in an airtight container until further use for biochemical and antioxidant assays.
Preparation of Extract Stock Solution
A stock solution of the Beta vulgaris ethanol extract was prepared by dissolving a measured quantity of the dried extract in dimethyl sulfoxide (DMSO). Working concentrations were subsequently prepared by serial dilution with culture medium (DMEM), ensuring that the final concentration of DMSO in all treatment wells did not exceed 0.1%, a level considered non-toxic to HepG2 cells.
2.3. Antioxidant Radical Scavenging Assays
2.3.1. DPPH Radical Scavenging Assay
The DPPH radical scavenging activity of the
Beta vulgaris ethanol extract was measured using the method described by Brand-Williams
et al.
with some modifications.
Fresh DPPH solution (0.1 mM) was prepared in methanol and mixed with different extract concentrations (100–600 µg/mL). Also, known concentrations of a standard antioxidant (ascorbic acid) was prepared too. For blank, DPPH was mixed with methanol only. The reaction mixtures were incubated in the dark for 30 minutes at room temperature. Absorbance was measured at 517 nm using a spectrophotometer. The % DPPH radical scavenging activity was calculated using equation (
1).
(1)
The IC50 value (concentration of extract that scavenges 50% of DPPH radicals) was also determined using a regression line plotted between percentage inhibition and extract concentration.
2.3.2. Hydrogen Peroxide (H2O2) Scavenging Assay
The hydrogen peroxide (H
2O
2) radical scavenging activity of the
Beta vulgaris ethanol extract was determined using the method described by Ruch
et al.
| [26] | Ruch, R. J., Cheng, S. J., & Klaunig, J. E. (1989). Prevention of cytotoxicity and inhibition of intercellular communication by antioxidant catechins isolated from Chinese green tea. Carcinogenesis, 10(6), 1003-1008.
https://doi.org/10.1093/carcin/10.6.1003 |
[26]
with some minor modifications. A 40 mM H
2O
2 solution was prepared in phosphate buffer (pH 7.4) and mixed with varying concentration of the extract (100–600 µg/mL). Also, known concentrations of a standard natural antioxidant (ascorbic acid) was prepared too. For the blank, H
2O
2 was mixed with solvent only. The mixtures were incubated for 10 minutes at room temperature. The absorbance was afterwards measured at 230nm with a spectrophotometer and recorded. The H
2O
2 scavenging ability was calculated using equation (
2).
(2)
The IC50 value (concentration of extract that scavenges 50% of H2O2 radicals) was also determined using a regression line plotted between percentage inhibition and extract concentration.
2.4. Cell Culture and Maintenance
Human hepatocellular carcinoma (HepG2) cells were obtained from a certified cell repository National Centre for Cell Sciences (NCCS, Pune, India) and maintained under standard culture conditions. Cells were grown in Dulbecco’s Modified Eagle Medium (DMEM) supplemented with 10% fetal bovine serum (FBS) and 1% penicillin–streptomycin to prevent microbial contamination. Cultures were incubated at 37°C in a humidified atmosphere containing 5% CO2. The growth medium was replaced every 2–3 days, and cells were subcultured at approximately 70–80% confluence using trypsin–EDTA.
2.4.1. Seeding of Cells for Experimental Assays
For experimental analysis, HepG2 cells were seeded into 96-well plates (1 × 10⁴ cells/well) for cell viability assays and for 6-well plates (2 × 10⁵ cells/well) for biochemical oxidative stress assays. The cells were allowed to attach overnight before treatment.
2.4.2. Induction of Oxidative Stress and Cytotoxic Model
Oxidative stress was induced by exposing HepG2 cells to hydrogen peroxide. To establish the oxidative injury model, cells were treated with different concentrations of H2O2 (50–500 µM) for 24 hours. The concentration causing approximately 50% reduction in viability (IC₅₀) was selected for subsequent experiments.
2.4.3. Experimental Design and Treatment Groups
Cells were randomly divided into eight groups (8), the Normal Control Group (the untreated HepG2 cells maintained in culture medium only), Oxidative Stress Control Group (cells exposed to H2O2 only), Extract Pre-treatment Groups which are cells pretreated with Beta vulgaris ethanol extracts at different concentrations (200, 400, 600, 800 and 1000 µg/ml) for 24 hours followed by H2O2 exposure, and Positive Control Group (cells were treated with a standard drug Silymarin prior to H2O2 induction).
Table 1. Treatment Protocol.
S/No. | Groups | ID |
1 | Normal control | Untreated Control |
2 | Hydrogen Peroxide (H2O2) only | Negative Control |
3 | Hydrogen Peroxide (H2O2) + 150mg/ml silymarin | Positive control |
4 | Test 1 (H2O2+ 200mg/ml) | HH200 |
5 | Test 2 (H2O2+ 400mg/ml) | HH400 |
6 | Test 3 (H2O2+ 600mg/ml) | HH600 |
7 | Test 4 (H2O2+ 800mg/ml) | HH800 |
8 | Test 5 (H2O2+ 1000mg/ml) | HH1000 |
2.5. Cell Viability Assay (MTT Assay)
Cell viability was assessed using the MTT colorimetric assay. The HepG2 cells were seeded into 96-well plates and treated according to experimental grouping. Following treatment, MTT solution (0.5 mg/mL) was added to each well and incubated for 4 hours at 37°C. The resulting formazan crystals were dissolved in dimethyl sulfoxide (DMSO), and absorbance was measured at 570 nm using a microplate reader. The cell viability was expressed as a percentage relative to the untreated control group.
2.6. Preparation of Cell Lysates for Biochemical Assays
Following treatment, cells were harvested and washed twice with cold phosphate-buffered saline (PBS). Cells were lysed using ice-cold phosphate buffer and centrifuged at 10,000 rpm for 15 min at 4°C. The resulting supernatant was collected and used for the determination of oxidative stress biomarkers and antioxidant enzyme activities.
2.7. Biochemical Assessment of Oxidative Stress Biomarkers
2.7.1. Lipid Peroxidation Assay (MDA Determination)
Lipid peroxidation was evaluated by measuring malondialdehyde (MDA) levels using the thiobarbituric acid reactive substances (TBARS) method as described by
. The cell lysates were mixed with thiobarbituric acid reagent and heated in a boiling water bath. After cooling, absorbance was measured at 532 nm. MDA concentration was expressed as nmol/mg protein.
2.7.2. Reduced Glutathione (GSH) Assay
Reduced glutathione (GSH) levels were determined using Ellman’s reagent (DTNB) as described by Ellman
. Cell lysates were mixed with DTNB and absorbance was read at 412 nm. Results were expressed as µmol/mg protein. The GSH levels were calculated using the standard calibration curve - a plot of the GSH standard absorbances vs concentration of the GSH in µM.
2.7.3. Determination of Superoxide Dismutase (SOD) Activity
SOD activity was determined using the pyrogallol autooxidation method as described by Marklund & Marklund
| [22] | Marklund, S., & Marklund, G. (1974). Involvement of the superoxide anion radical in the autoxidation of pyrogallol and a convenient assay for superoxide dismutase. European Journal of Biochemistry, 47(3), 469-474.
https://doi.org/10.1111/j.1432-1033.1974.tb03714.x |
[22]
. The assay is based on the ability of SOD to inhibit the autoxidation of pyrogallol in alkaline Tris–HCl buffer (pH 8.2) that is containing 1mM EDTA. One unit of SOD activity was defined as the amount of enzyme causing 50% inhibition of pyrogallol oxidation, and results were expressed as U/mg protein.
2.7.4. Catalase (CAT) Activity
Catalase activity was determined by monitoring the decomposition rate of hydrogen peroxide at 240 nm as described by Aebi
CAT activity was expressed as µmol H
2O
2 decomposed/min/mg protein.
2.8. Statistical Analysis
All experiments were performed in triplicate (n = 3) and data were analyzed using SPSS version 20. Data were expressed as mean ± standard error of the mean (SEM). Statistical analysis was conducted using one-way analysis of variance (ANOVA) followed by Tukey’s post hoc test. Differences were considered statistically significant at p < 0.05. Graphical representation and statistical analyses were performed using Microsoft Excel. Calibration curves were constructed using linear regression analysis, and the coefficient of determination (R²) was reported.
3. Results
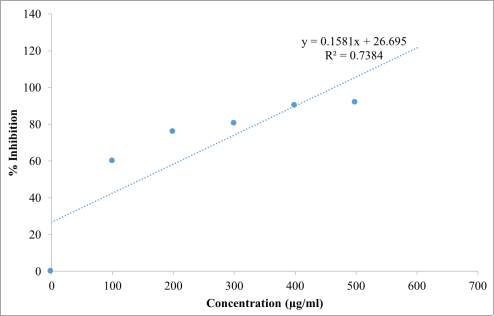
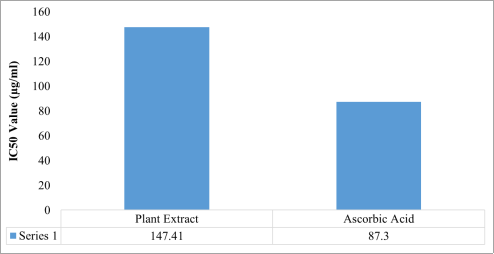
The results of the DPPH radical scavenging assay (
Table 2;
Figures 1-3) demonstrated that the
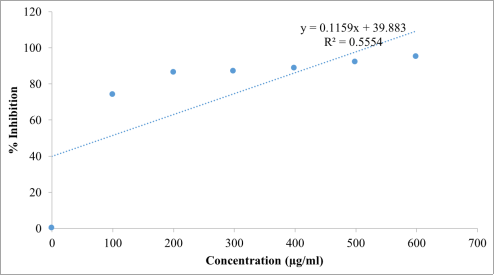
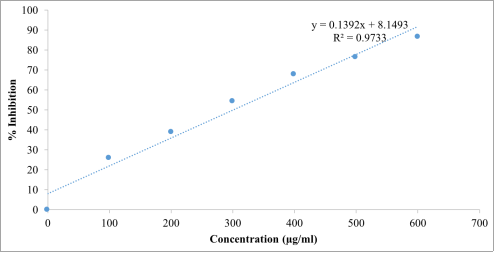
Beta vulgaris ethanol extract exhibited a progressive increase in antioxidant activity with increasing concentration, indicating a strong concentration-dependent antioxidant response. The highest scavenging activity was recorded at 600 µg/mL, with 93.14% inhibition, which was comparable to the activity of ascorbic acid (95.10%) at the same concentration. Regression analysis further confirmed a linear relationship between concentration and radical scavenging activity for both the extract and the standard antioxidant. The IC₅₀ value of the extract was 147.41 µg/mL, which, although higher than that of ascorbic acid (87.30 µg/mL), suggests that the extract possesses substantial free radical scavenging potential. Similarly, the hydrogen peroxide scavenging assay results (
Table 3;
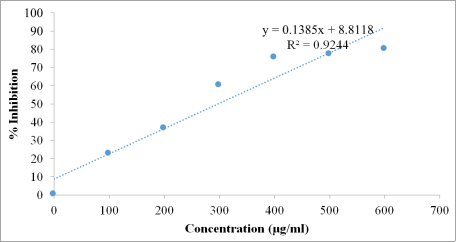
Figures 4-6) revealed that the
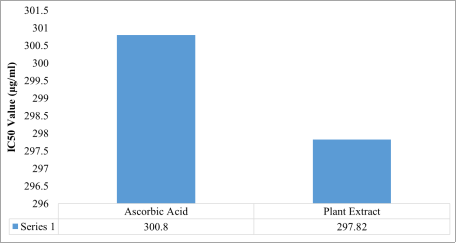
Beta vulgaris ethanol extract displayed a concentration-dependent increase in scavenging activity. The extract showed comparable effectiveness to ascorbic acid across most tested concentrations. Regression analyses confirmed strong linear relationships between concentration and scavenging activity for both treatments. Notably, the IC₅₀ value of the extract (297.82 µg/mL) was slightly lower than that of ascorbic acid (300.80 µg/mL), indicating that the extract exhibited marginally superior hydrogen peroxide scavenging efficiency.
Table 2. DPPH Radical Scavenging Test showing % Inhibition of Ascorbic Acid and Beta vulgaris Ethanol Extract.
S/No. | Concentration (µg/ml) | % Inhibition of Beta vulgaris Ethanol Extract | % Inhibition of Ascorbic Acid |
2 | 100 | 59.80 | 73.85 |
3 | 200 | 75.50 | 85.95 |
4 | 300 | 80.40 | 86.93 |
5 | 400 | 90.10 | 88.56 |
6 | 500 | 91.50 | 92.10 |
7 | 600 | 93.14 | 95.10 |
Figure 1. Regression line and equation of different concentration of Beta vulgaris ethanol extract with DPPH solution in the antioxidant activity test.
Figure 2. Regression line and equation of different concentration of ascorbic acid with DPPH solution in the antioxidant activity test.
Figure 3. The IC50 values for the Beta vulgaris ethanol extract and ascorbic acid for DPPH radical scavenging activity.
Table 3. Hydrogen Peroxide Radical Scavenging Test showing % Inhibition of Ascorbic Acid and Beta vulgaris Ethanol Extract.
S/No. | Concentration (µg/ml) | % Inhibition of Beta vulgaris Ethanol Extract | % Inhibition of Ascorbic Acid |
2 | 100 | 22.88 | 25.75 |
3 | 200 | 36.60 | 38.82 |
4 | 300 | 60.47 | 54.25 |
5 | 400 | 75.16 | 67.58 |
6 | 500 | 77.38 | 76.47 |
7 | 600 | 80.10 | 86.54 |
Figure 4. Regression line and regression equation of different concentration of ascorbic acid with hydrogen peroxide solution in the antioxidant activity test.
Figure 5. Regression line and regression equation of different concentration of Beta vulgaris ethanol extract with hydrogen peroxide solution in the antioxidant activity test.
Figure 6. IC50 values for hydrogen peroxide radical scavenging activity for the Beta vulgaris ethanol extract and ascorbic acid.
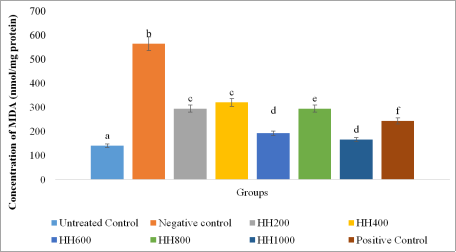
The malondialdehyde (MDA) levels in treated HepG2 cells are presented in
Figure 7. The results showed a significant increase (
p < 0.05) in MDA concentration in the negative control (H
2O
2-treated group) compared with the untreated control, confirming successful induction of lipid peroxidation and oxidative stress. Treatment with the
Beta vulgaris ethanol extract at different concentrations (HH200–HH1000) produced variable reductions in MDA levels, indicating protective effects against oxidative membrane damage. The optimal inhibitory effect was observed at HH600 and HH1000 which was significantly different (
p > 0.05) from that of the positive control, silymarin.
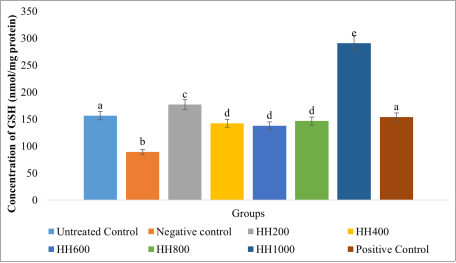
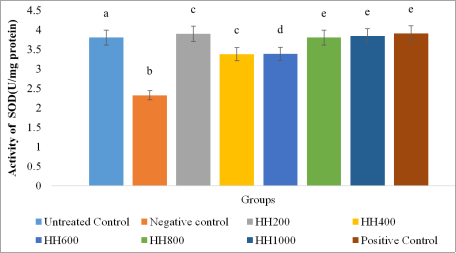
The results for intracellular antioxidant markers, reduced glutathione (GSH), catalase (CAT), and superoxide dismutase (SOD), are presented in
Figures 8-10. The untreated control group represents baseline antioxidant status in HepG2 cells under normal conditions. In the negative control group exposed to H
2O
2 alone (
Figure 8), GSH levels decreased significantly (
p < 0.05) to 89.05 nmol/mg protein, indicating depletion of intracellular glutathione during detoxification of reactive oxygen species. This further confirms the successful induction of oxidative stress in the cells. Pretreatment with the plant extract (HH200–HH1000) significantly improved GSH levels relative to the negative control. At HH200, GSH increased significantly (
p < 0.05) to 176.89 nmol/mg protein, suggesting a protective effect at this concentration, possibly through stimulation of GSH synthesis or reduced consumption. At intermediate concentrations (HH400–HH800), GSH levels ranged between 142.09 and 146.49 nmol/mg protein, remaining significantly higher than the negative control group but slightly lower than the untreated control, indicating moderate protection. The highest concentration (HH1000) produced a marked increase in GSH to 290.41 nmol/mg protein, nearly double the untreated level, suggesting a strong antioxidant response potentially mediated by enhanced glutathione biosynthesis and efficient ROS scavenging. The positive control (silymarin) produced a GSH level of 153.92 nmol/mg protein, indicating mild but consistent protection in line with its established hepatoprotective activity.
Also, H
2O
2 treatment significantly decreased SOD activity from 3.80 to 2.32 U/mg protein (
p < 0.05), indicating oxidative suppression of superoxide detoxification capacity (
Figure 9). Extract treatment resulted in concentration-dependent recovery of SOD activity. Low (HH200) and high concentrations (HH800–HH1000) almost completely restored SOD levels, with values significantly different (
p > 0.05) from the untreated control group and not significantly different (
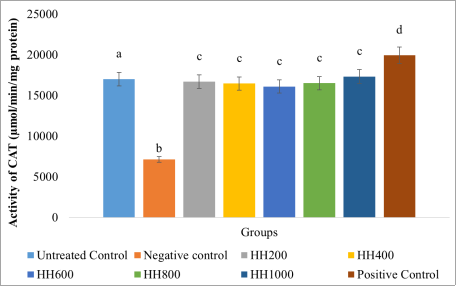
p > 0.05) from the positive control group. Moderate concentrations (HH400–HH600) showed partial restoration. Similarly, hydrogen peroxide exposure also resulted in a significant reduction (
p < 0.05) in catalase activity compared to untreated cells, confirming oxidative stress-induced impairment of enzymatic antioxidant defense (
Figure 10). Treatment with the extract restored CAT activity, with the extract concentrations producing elevated enzyme levels with values not significantly different (
p > 0.05) across the extract treated groups. These effects were comparable, though slightly lower, than those observed with silymarin. The results indicated that
Beta vulgaris ethanol extract significantly enhanced both non-enzymatic (GSH) and enzymatic (CAT and SOD) antioxidant defenses, demonstrating strong cytoprotective activity against H
2O
2-induced oxidative damage. The antioxidant efficacy of the extract was comparable to that of the standard hepatoprotective agent, silymarin.
Figure 7. Malondialdehyde (MDA) levels in the HepG2 cells.
Values are represented as (Mean± S. E. M) Means with different superscript (a-f) are significantly different (Tukey hsd, p<0.05). Superscript letters (a - f) compares the malondialdehyde levels across the experimental groups.
Figure 8. Reduced glutathione (GSH) levels in the cells.
Values are represented as (Mean ± S. E. M). Means with different superscript (a-e) are significantly different (Tukey hsd, p<0.05). Superscript letters (a - e) compares the reduced glutathione levels across the experimental groups.
Figure 9. SOD Activity in Cells.
Values are represented as (Mean±S. E. M). Means with different superscript (a-e) are significantly different (Tukey hsd, p<0.05). Superscript letters (a-e) compares superoxide dismutase activity across the experimental groups.
Figure 10. Catalase (CAT) activity in the cells.
Values are represented as (Mean±S. E. M) Means with different superscript (a-d) are significantly different (Tukey hsd, p<0.05). Superscript letters (a-d) compares catalase activity across the experimental groups.
4. Discussion
From the analysis conducted, the results of the scavenging assays indicate that the
Beta vulgaris ethanol extract exhibits significant antioxidant activity, demonstrated by the concentration-dependent increase in its ability to neutralize free radicals aligning with the concept that extracts rich in phytochemicals tend to donate electrons or hydrogen atoms more effectively at higher concentrations, thereby enhancing the reduction of free radicals
| [6] | Brand-Williams, W., Cuvelier, M. E., & Berset, C. (1995). Use of a free radical method to evaluate antioxidant activity. LWT - Food Science and Technology, 28(1), 25-30.
https://doi.org/10.1016/S0023-6438(95)80008-5 |
| [14] | Halliwell, B., & Gutteridge, J. M. C. (2015). Free radicals in biology and medicine (5th ed.). Oxford University Press. |
| [23] | Molyneux, P. (2004). The use of the stable free radical diphenylpicrylhydrazyl (DPPH) for estimating antioxidant activity. Songklanakarin Journal of Science and Technology, 26(2), 211-219. |
| [26] | Ruch, R. J., Cheng, S. J., & Klaunig, J. E. (1989). Prevention of cytotoxicity and inhibition of intercellular communication by antioxidant catechins isolated from Chinese green tea. Carcinogenesis, 10(6), 1003-1008.
https://doi.org/10.1093/carcin/10.6.1003 |
[6, 14, 23, 26]
. The DPPH radical scavenging assay evaluates the capacity of compounds to donate electrons or hydrogen atoms, thereby neutralizing free radicals
| [23] | Molyneux, P. (2004). The use of the stable free radical diphenylpicrylhydrazyl (DPPH) for estimating antioxidant activity. Songklanakarin Journal of Science and Technology, 26(2), 211-219. |
[23]
. Similarly, the hydrogen peroxide scavenging assay determines the ability of extracts to detoxify H
2O
2, a reactive oxygen species capable of crossing membranes and forming highly reactive hydroxyl radicals
| [26] | Ruch, R. J., Cheng, S. J., & Klaunig, J. E. (1989). Prevention of cytotoxicity and inhibition of intercellular communication by antioxidant catechins isolated from Chinese green tea. Carcinogenesis, 10(6), 1003-1008.
https://doi.org/10.1093/carcin/10.6.1003 |
[26]
. The observed antioxidant effect in both DPPH and H
2O
2 scavenging assays is likely due to the presence of bioactive constituents naturally found in
Beta vulgaris, consistent with previous studies attributing the strong scavenging ability of
Beta vulgaris to its richness in betalains, phenolics, and flavonoids, and supporting its use as a source of bioactive compounds with health-promoting and disease-preventive properties
| [8] | Chhikara, N., Kushwaha, K., Sharma, P., Gat, Y., & Panghal, A. (2019). Bioactive compounds of beetroot and utilization in food processing industry: A critical review. Food Chemistry, 272, 192-200. https://doi.org/10.1016/j.foodchem.2018.08.022 |
| [9] | Clifford, T., Howatson, G., West, D. J., & Stevenson, E. J. (2015). The potential benefits of red beetroot supplementation in health and disease. Nutrients, 7(4), 2801-2822.
https://doi.org/10.3390/nu7042801 |
| [13] | Georgiev, V. G., Weber, J., Kneschke, E. M., Denev, P. N., Bley, T., & Pavlov, A. I. (2010). Antioxidant activity and phenolic content of betalain extracts from intact plants and hairy root cultures of the red beetroot Beta vulgaris cv. Detroid dark red. Plant Foods for Human Nutrition, 65(2), 105-111.
https://doi.org/10.1007/s11130-010-0156-6 |
| [19] | Kujala, T. S., Vienola, M. S., Klika, K. D., Loponen, J. M., & Pihlaja, K. (2002). Betalain and phenolic compositions of four beetroot (Beta vulgaris) cultivars. European Food Research and Technology, 214(6), 505-510.
https://doi.org/10.1007/s00217-001-0478-6 |
| [23] | Molyneux, P. (2004). The use of the stable free radical diphenylpicrylhydrazyl (DPPH) for estimating antioxidant activity. Songklanakarin Journal of Science and Technology, 26(2), 211-219. |
| [25] | Ojo, O. A., Adeyemo, T. R., Iyobhebhe, M., Adams, M. D., Asaleye, R. M., Evbuomwan, I. O., Abdurrahman, J., Maduakolam-Aniobi, T. C., Nwonuma, C. O., Odesanmi, O. E., & Ojo, A. B. (2024). Beta vulgaris L. beetroot protects against iron-induced liver injury by restoring antioxidant pathways and regulating cellular functions. Scientific reports, 14(1), 25205.
https://doi.org/10.1038/s41598-024-77503-6 |
[8, 9, 13, 19, 23, 25]
. Similar patterns have also been noted in other research, where the synergistic interaction of multiple phytochemicals in
Beta vulgaris extracts led to heightened antioxidant activity
| [8] | Chhikara, N., Kushwaha, K., Sharma, P., Gat, Y., & Panghal, A. (2019). Bioactive compounds of beetroot and utilization in food processing industry: A critical review. Food Chemistry, 272, 192-200. https://doi.org/10.1016/j.foodchem.2018.08.022 |
[8]
. Additionally, the
Beta vulgaris ethanol extract’s performance relative to ascorbic acid is consistent with existing literature, which emphasizes that crude plant extracts often exhibit slightly higher IC₅₀ values than pure standards but still retain substantial antioxidant activity due to the synergistic interactions among multiple phytochemicals
| [21] | Li, S., Tan, H. Y., Wang, N., Zhang, Z. J., Lao, L., Wong, C. W., & Feng, Y. (2015). The role of oxidative stress and antioxidants in liver diseases. International Journal of Molecular Sciences, 16(11), 26087-26124.
https://doi.org/10.3390/ijms161125942 |
| [25] | Ojo, O. A., Adeyemo, T. R., Iyobhebhe, M., Adams, M. D., Asaleye, R. M., Evbuomwan, I. O., Abdurrahman, J., Maduakolam-Aniobi, T. C., Nwonuma, C. O., Odesanmi, O. E., & Ojo, A. B. (2024). Beta vulgaris L. beetroot protects against iron-induced liver injury by restoring antioxidant pathways and regulating cellular functions. Scientific reports, 14(1), 25205.
https://doi.org/10.1038/s41598-024-77503-6 |
[21, 25]
.
Hydrogen peroxide is well documented to trigger oxidative stress–mediated responses in liver cells, resulting in the elevation of intracellular reactive oxygen species (ROS), leading to lipid peroxidation, depletion of glutathione (GSH), and impairment of antioxidant enzymes such as superoxide dismutase (SOD), catalase (CAT), and glutathione peroxidase (GPx)
| [16] | Je, J. Y. & Lee, D. B. (2015). Nelumbo nucifera leaves protect hydrogen peroxide-induced hepatic damage via antioxidant enzymes and HO-1/Nrf2 activation. Food and Function, 6(6), 1911-1918. https://doi.org/10.1039/c5fo00201j |
[16]
. To counteract reactive oxygen species (ROS), cells rely on a sophisticated antioxidant defense network that maintains redox homeostasis
| [29] | Váli, L., Stefanovits-Bányai, E., Szentmihályi, K., Fébel, H., Sárdi, E., Lugasi, A., Kocsis, I., & Blázovics, A. (2007). Liver-protecting effects of table beet (Beta vulgaris var. rubra) during ischemia-reperfusion. Nutrition, 23(2), 172-178.
https://doi.org/10.1016/j.nut.2006.11.004 |
[29]
. This system includes enzymatic antioxidants - SOD, CAT, and peroxidases (PRx), and non-enzymatic molecules such as GSH, vitamins C and E, carotenoids, and polyphenolic compounds derived from plants
| [5] | Birben, E., Sahiner, U. M., Sackesen, C., Erzurum, S., & Kalayci, O. (2012). Oxidative stress and antioxidant defense. World Allergy Organization Journal, 5(1), 9-19.
https://doi.org/10.1097/WOX.0b013e3182439613 |
| [29] | Váli, L., Stefanovits-Bányai, E., Szentmihályi, K., Fébel, H., Sárdi, E., Lugasi, A., Kocsis, I., & Blázovics, A. (2007). Liver-protecting effects of table beet (Beta vulgaris var. rubra) during ischemia-reperfusion. Nutrition, 23(2), 172-178.
https://doi.org/10.1016/j.nut.2006.11.004 |
[5, 29]
.
In the
in vitro cell based antioxidant assays, the observed decrease and increase in antioxidant levels in the negative control group of the experimental groups confirms that hydrogen peroxide successfully induced oxidative stress in HepG2 cells
| [14] | Halliwell, B., & Gutteridge, J. M. C. (2015). Free radicals in biology and medicine (5th ed.). Oxford University Press. |
[14]
. Treatment with
Beta vulgaris ethanol extract markedly restored GSH levels, significantly reduced MDA levels and formation, counteracted CAT suppression; progressively improved CAT activity at all tested concentrations, and significantly elevated SOD levels. This indicates significant antioxidant and hepatoprotective effects.
These findings are in agreement with previous reports and studies highlighting the richness of
Beta vulgaris in bioactive compounds capable of promoting GSH synthesis, improving CAT activity, reducing MDA levels effectively preventing lipid peroxidation, substantially restoring SOD activity, maintaining cellular redox equilibrium, and the protective activity likely due to the compounds stimulating antioxidant pathways and modulating enzyme expression, functioning as chain-breaking antioxidants, scavenging ROS and stabilizing lipid radicals through the neutralization of hydroxyl and peroxyl radicals
| [2] | Agarwal, M., Srivastava, V. K., Saxena, K. K., & Kumar, A. (2006). Hepatoprotective activity of Beta vulgaris against CCl4-induced hepatic injury in rats. Fitoterapia, 77(2), 91-93.
https://doi.org/10.1016/j.fitote.2005.11.004 |
| [3] | Al-Harbi, L. N., Alshammari, G. M., Al-Dossari, A. M., Subash-Babu, P., Binobead, M. A., Alhussain, M. H., AlSedairy, S. A., Al-Nouri, D. M., & Shamlan, G. (2021). Beta vulgaris L. (Beetroot) Methanolic Extract Prevents Hepatic Steatosis and Liver Damage in T2DM Rats by Hypoglycemic, Insulin-Sensitizing, Antioxidant Effects, and Upregulation of PPARα. Biology, 10(12), 1306. https://doi.org/10.3390/biology10121306 |
| [7] | Carlsen, M. H., Halvorsen, B. L., Holte, K., Bøhn, S. K., Dragland, S., Sampson, L., Willey, C., Senoo, H., Umezono, Y., Sanada, C., Barikmo, I., Berhe, N., Willett, W. C., Phillips, K. M., Jacobs, D. R., & Blomhoff, R. (2010). The total antioxidant content of more than 3100 foods, beverages, spices, herbs and supplements used worldwide. Nutrition Journal, 9(1), 3.
https://doi.org/10.1186/1475-2891-9-3 |
| [9] | Clifford, T., Howatson, G., West, D. J., & Stevenson, E. J. (2015). The potential benefits of red beetroot supplementation in health and disease. Nutrients, 7(4), 2801-2822.
https://doi.org/10.3390/nu7042801 |
| [12] | Esatbeyoglu, T., Wagner, A. E., Schini-Kerth, V. B., & Rimbach, G. (2015). Betanin - a food colorant with biological activity. Molecular Nutrition & Food Research, 59(1), 36-47.
https://doi.org/10.1002/mnfr.201400484 |
| [13] | Georgiev, V. G., Weber, J., Kneschke, E. M., Denev, P. N., Bley, T., & Pavlov, A. I. (2010). Antioxidant activity and phenolic content of betalain extracts from intact plants and hairy root cultures of the red beetroot Beta vulgaris cv. Detroid dark red. Plant Foods for Human Nutrition, 65(2), 105-111.
https://doi.org/10.1007/s11130-010-0156-6 |
| [19] | Kujala, T. S., Vienola, M. S., Klika, K. D., Loponen, J. M., & Pihlaja, K. (2002). Betalain and phenolic compositions of four beetroot (Beta vulgaris) cultivars. European Food Research and Technology, 214(6), 505-510.
https://doi.org/10.1007/s00217-001-0478-6 |
| [25] | Ojo, O. A., Adeyemo, T. R., Iyobhebhe, M., Adams, M. D., Asaleye, R. M., Evbuomwan, I. O., Abdurrahman, J., Maduakolam-Aniobi, T. C., Nwonuma, C. O., Odesanmi, O. E., & Ojo, A. B. (2024). Beta vulgaris L. beetroot protects against iron-induced liver injury by restoring antioxidant pathways and regulating cellular functions. Scientific reports, 14(1), 25205.
https://doi.org/10.1038/s41598-024-77503-6 |
| [28] | Sies, H., Berndt, C., & Jones, D. P. (2017). Oxidative stress. Annual Review of Biochemistry, 86, 715-748.
https://doi.org/10.1146/annurev-biochem-061516-045037 |
[2, 3, 7, 9, 12, 13, 19, 25, 28]
.