Acinetobacter baumannii is one of the most significant threats to global public health with a seemingly limitless capacity for antimicrobial resistance. Developing preventive vaccines to combat these pathogens can provide long-term protective immunity. This study was conducted to evaluate the specific protein against which the protective antibody is produced in serum and splenic cell culture supernatant after vaccination in mice. The vaccine used was a formaldehyde-inactivated whole-cell vaccine against multidrug resistant (MDR) A. baumannii. MDR A. baumannii, isolated from different samples, used in formaldehyde-inactivated vaccine and inoculated intradermally in experimental mice. Serum was collected from tail blood on days 13, 27 and 41 following 1st immunization. Two weeks after 3rd immunization, mice were challenged with live A. baumannii intraperitoneally and observed for 14 days. Sera from tail blood and mouse spleens were collected. MDR A. baumannii antigen was sonicated and Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis was performed. Protective antibodies that bind with antigens in serum and culture supernatant were evaluated by ELISA. Survival rates were observed to be 100% at 14 days post-challenge among the immunized mice. Serum from pre- and post-challenge immunizations showed considerably greater optical density (OD) values of IgG than control mice. Although antibodies are developed against several antigens of different molecular weights, the protein fraction of 30-40 kDa showed a higher OD value. Formalin-inactivated intradermal immunization with MDR A. baumannii produced protective antibodies in mice against proteins of specific molecular weights. Proteins of 30-40 kDa molecular weights could be used as vaccine candidates to prevent A. baumannii infection.
| Published in | International Journal of Immunology (Volume 13, Issue 1) |
| DOI | 10.11648/j.iji.20251301.11 |
| Page(s) | 1-8 |
| Creative Commons |
This is an Open Access article, distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution and reproduction in any medium or format, provided the original work is properly cited. |
| Copyright |
Copyright © The Author(s), 2025. Published by Science Publishing Group |
A. baumannii, Antibody, Bangladesh, Dhaka, Immunization, SDS-PAGE, Whole Cell Vaccine
Vaccine schedule | OD value range | Mean of negative control | Standard deviation | Cut off value |
|---|---|---|---|---|
1st inoculation | 0.517-0.735 | 0.131 | 0.008 | 0.147 |
2nd inoculation | 1.298-1.392 | 0.129 | 0.002 | 0.132 |
3rd inoculation | 1.519-1.930 | 0.129 | 0.227 | 0.142 |
Post challenge | 1.253-1.651 | 0.133 | 0.005 | 0.144 |
AEEC | Animal Experimentation Ethics Committee |
CFU | Colony Forming Unit |
ELISA | Enzyme-Linked Immunosorbent Assay |
ERC | Ethical Review Committee |
Icddr, b | International Center for Diarrheal Disease Research, Bangladesh |
ICU | Intensive Care Unit |
MDR | Multidrug Resistant |
OD | Optical Density |
PBS | Phosphate Buffered Saline |
PCR | Polymerase Chain Reaction |
RPMI | Roswell Park Memorial 1640 Medium |
SDS-PAGE | Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis |
TEMED | N, N, N’, N’-Tetramethylethylenediamine |
TSB | Tryptic Soy Broth |
WHO | World Health Organization |
| [1] | Singh R, Capalash N, Sharma P. Vaccine development to control the rising scourge of antibiotic-resistant Acinetobacter baumannii: a systematic review. 3 Biotech 2022; 12: 85. |
| [2] | Vázquez-López R, Solano-Gálvez SG, Juárez Vignon-Whaley JJ, Abello Vaamonde JA, Padró Alonzo LA, Rivera Reséndiz A, et al. Acinetobacter baumannii Resistance: A Real Challenge for Clinicians. Antibiotics 2020; 9: 205. |
| [3] | Perez F, Hujer AM, Hujer KM, Decker BK, Rather PN, Bonomo RA. Global Challenge of Multidrug-Resistant Acinetobacter baumannii. Antimicrobial Agents and Chemotherapy 2007; 51: 3471-84. |
| [4] | Howard A, O’Donoghue M, Feeney A, Sleator RD. Acinetobacter baumannii: An emerging opportunistic pathogen. Virulence 2012; 3: 243-50. |
| [5] | Shan W, Kan J, Cai X, Yin M. Insights into mucoid Acinetobacter baumannii: A review of microbiological characteristics, virulence, and pathogenic mechanisms in a threatening nosocomial pathogen. Microbiological Research 2022; 261: 127057. |
| [6] | Neshani A, Sedighian H, Mirhosseini SA, Ghazvini K, Zare H, Jahangiri A. Antimicrobial peptides as a promising treatment option against Acinetobacter baumannii infections. Microbial Pathogenesis 2020; 146: 104238. |
| [7] | Fan Y, Mu Y, Lu L, Tian Y, Yuan F, Zhou B, et al. Hydrogen peroxide-inactivated bacteria induces potent humoral and cellular immune responses and releases nucleic acids. International Immunopharmacology 2019; 69: 389-97. |
| [8] | Dennehy R, McClean S. Immunoproteomics: The Key to Discovery of New Vaccine Antigens Against Bacterial Respiratory Infections. CPPS 2012; 13: 807-15. |
| [9] | Takemori N, Takemori A. In-depth structural proteomics integrating mass spectrometry and polyacrylamide gel electrophoresis. Front Anal Sci 2023; 2. |
| [10] | Kawser Z, Shamsuzzaman SM. Intradermal Immunization with Heat-Killed Klebsiella pneumoniae Leading to the Production of Protective Immunoglobulin G in BALB/c Mice. International Journal of Applied and Basic Medical Research 2021; 11: 160. |
| [11] | Agallou M, Karagouni E. Detection of Antigen-specific T cells in Spleens of Vaccinated Mice Applying 3[H]-Thymidine Incorporation Assay and Luminex Multiple Cytokine Analysis Technology. BIO-PROTOCOL 2019; 9. |
| [12] |
Strober W. Trypan Blue Exclusion Test of Cell Viability - Strober - 2015 - Current Protocols in Immunology - Wiley Online Library 2015.
https://currentprotocols.onlinelibrary.wiley.com/doi/abs/10.1002/0471142735.ima03bs111 (accessed November 11, 2024). |
| [13] | Green MR, Sambrook J. Molecular cloning A Laboratory Manual. 4th ed. Cold Spring Harbor Laboratory Press: 2012, 1599-1610. |
| [14] |
ThermoFisher Scientific. Extract proteins from polyacrylamide gels. Available from:
https://assets.thermofisher.com/TFS-Assets/BID/Technical-Notes/extract-proteins-polyacrylamide-gels-tech-tip.pdf (accessed 2 February 2025). |
| [15] | McConnell MJ, Pachón J. Active and passive immunization against Acinetobacter baumannii using an inactivated whole cell vaccine. Vaccine 2010; 29: 1-5. |
| [16] | Khan MA, Allemailem KS, Maswadeh H, Younus H. Glycosphingolipids (GSLs) from Sphingomonas paucimobilis Increase the Efficacy of Liposome-Based Nanovaccine against Acinetobacter baumannii-Associated Pneumonia in Immunocompetent and Immunocompromised Mice. Molecules 2022; 27: 7790. |
| [17] | Takahashi K, Hanamura Y, Tokunoh N, Kassai K, Matsunishi M, Watanabe S, et al. Protective effects of oral immunization with formalin-inactivated whole-cell Citrobacter rodentium on Citrobacter rodentium infection in mice. Journal of Microbiological Methods 2019; 159: 62-8. |
| [18] | Sheweita SA, Batah AM, Ghazy AA, Hussein A, Amara AA. A new strain of Acinetobacter baumannii and characterization of its ghost as a candidate vaccine. Journal of Infection and Public Health 2019; 12: 831-42. |
APA Style
Hasan, K., Shamsuzzaman, S., Rashid, M. H. U., Parveen, F. (2025). Antibody Responses Against Proteins of Different Molecular Weight Following Inoculation of Formalin Inactivated Acinetobacter baumannii in Mice. International Journal of Immunology, 13(1), 1-8. https://doi.org/10.11648/j.iji.20251301.11
ACS Style
Hasan, K.; Shamsuzzaman, S.; Rashid, M. H. U.; Parveen, F. Antibody Responses Against Proteins of Different Molecular Weight Following Inoculation of Formalin Inactivated Acinetobacter baumannii in Mice. Int. J. Immunol. 2025, 13(1), 1-8. doi: 10.11648/j.iji.20251301.11
AMA Style
Hasan K, Shamsuzzaman S, Rashid MHU, Parveen F. Antibody Responses Against Proteins of Different Molecular Weight Following Inoculation of Formalin Inactivated Acinetobacter baumannii in Mice. Int J Immunol. 2025;13(1):1-8. doi: 10.11648/j.iji.20251301.11
@article{10.11648/j.iji.20251301.11,
author = {Kohinur Hasan and SM Shamsuzzaman and Mohammad Harun Ur Rashid and Farhana Parveen},
title = {Antibody Responses Against Proteins of Different Molecular Weight Following Inoculation of Formalin Inactivated Acinetobacter baumannii in Mice
},
journal = {International Journal of Immunology},
volume = {13},
number = {1},
pages = {1-8},
doi = {10.11648/j.iji.20251301.11},
url = {https://doi.org/10.11648/j.iji.20251301.11},
eprint = {https://article.sciencepublishinggroup.com/pdf/10.11648.j.iji.20251301.11},
abstract = {Acinetobacter baumannii is one of the most significant threats to global public health with a seemingly limitless capacity for antimicrobial resistance. Developing preventive vaccines to combat these pathogens can provide long-term protective immunity. This study was conducted to evaluate the specific protein against which the protective antibody is produced in serum and splenic cell culture supernatant after vaccination in mice. The vaccine used was a formaldehyde-inactivated whole-cell vaccine against multidrug resistant (MDR) A. baumannii. MDR A. baumannii, isolated from different samples, used in formaldehyde-inactivated vaccine and inoculated intradermally in experimental mice. Serum was collected from tail blood on days 13, 27 and 41 following 1st immunization. Two weeks after 3rd immunization, mice were challenged with live A. baumannii intraperitoneally and observed for 14 days. Sera from tail blood and mouse spleens were collected. MDR A. baumannii antigen was sonicated and Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis was performed. Protective antibodies that bind with antigens in serum and culture supernatant were evaluated by ELISA. Survival rates were observed to be 100% at 14 days post-challenge among the immunized mice. Serum from pre- and post-challenge immunizations showed considerably greater optical density (OD) values of IgG than control mice. Although antibodies are developed against several antigens of different molecular weights, the protein fraction of 30-40 kDa showed a higher OD value. Formalin-inactivated intradermal immunization with MDR A. baumannii produced protective antibodies in mice against proteins of specific molecular weights. Proteins of 30-40 kDa molecular weights could be used as vaccine candidates to prevent A. baumannii infection.
},
year = {2025}
}
TY - JOUR T1 - Antibody Responses Against Proteins of Different Molecular Weight Following Inoculation of Formalin Inactivated Acinetobacter baumannii in Mice AU - Kohinur Hasan AU - SM Shamsuzzaman AU - Mohammad Harun Ur Rashid AU - Farhana Parveen Y1 - 2025/03/31 PY - 2025 N1 - https://doi.org/10.11648/j.iji.20251301.11 DO - 10.11648/j.iji.20251301.11 T2 - International Journal of Immunology JF - International Journal of Immunology JO - International Journal of Immunology SP - 1 EP - 8 PB - Science Publishing Group SN - 2329-1753 UR - https://doi.org/10.11648/j.iji.20251301.11 AB - Acinetobacter baumannii is one of the most significant threats to global public health with a seemingly limitless capacity for antimicrobial resistance. Developing preventive vaccines to combat these pathogens can provide long-term protective immunity. This study was conducted to evaluate the specific protein against which the protective antibody is produced in serum and splenic cell culture supernatant after vaccination in mice. The vaccine used was a formaldehyde-inactivated whole-cell vaccine against multidrug resistant (MDR) A. baumannii. MDR A. baumannii, isolated from different samples, used in formaldehyde-inactivated vaccine and inoculated intradermally in experimental mice. Serum was collected from tail blood on days 13, 27 and 41 following 1st immunization. Two weeks after 3rd immunization, mice were challenged with live A. baumannii intraperitoneally and observed for 14 days. Sera from tail blood and mouse spleens were collected. MDR A. baumannii antigen was sonicated and Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis was performed. Protective antibodies that bind with antigens in serum and culture supernatant were evaluated by ELISA. Survival rates were observed to be 100% at 14 days post-challenge among the immunized mice. Serum from pre- and post-challenge immunizations showed considerably greater optical density (OD) values of IgG than control mice. Although antibodies are developed against several antigens of different molecular weights, the protein fraction of 30-40 kDa showed a higher OD value. Formalin-inactivated intradermal immunization with MDR A. baumannii produced protective antibodies in mice against proteins of specific molecular weights. Proteins of 30-40 kDa molecular weights could be used as vaccine candidates to prevent A. baumannii infection. VL - 13 IS - 1 ER -
Department of Microbiology, Dhaka Medical College, Dhaka, Bangladesh
Department of Microbiology, Dhaka Medical College, Dhaka, Bangladesh
Department of Intensive Care Unit, National Institute of Traumatology and Orthopedic Rehabilitation, Dhaka, Bangladesh
Department of Microbiology, Dhaka Medical College, Dhaka, Bangladesh
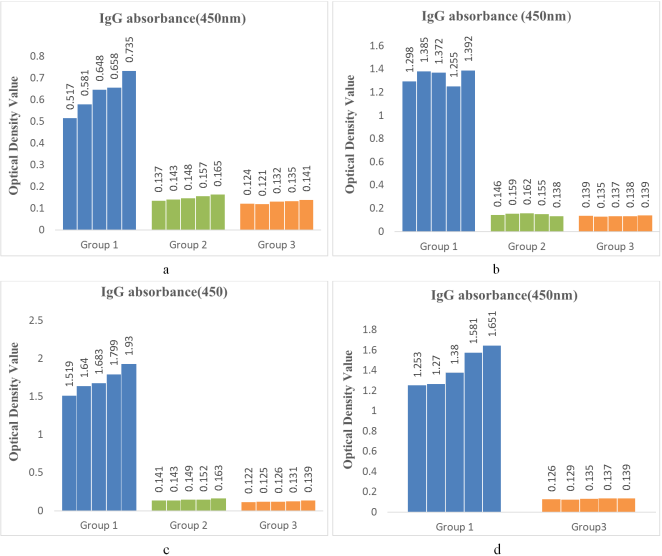
Figure 1. Optical density (OD) of immunized and unimmunized control serum samples after the first (a), second (b), third (c) inoculation, and post-challenge (d) by ELISA. Here, the OD values of IgG in serum samples collected on the 13th day following the first immunization showed that Group I had OD values above the cutoff value of 0.147 (a). On the 27th day following the first immunization, Group I had OD values above the cutoff value of 0.132 (b). On the 41st day following the first immunization, Group I had OD values above the cutoff value of 0.142 (c). After the lethal challenge, Group II mice did not survive, while Group I had OD values above the cutoff value of 0.144 (d). Here, Group I = immunized mice, Group II = unimmunized mice with PBS inoculation, and Group III = unimmunized and uninoculated mice. The cutoff values are mentioned in Table 1.
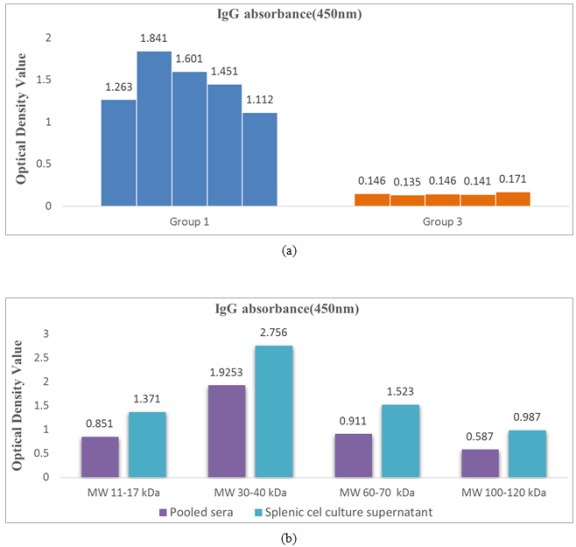
Figure 2. (a) IgG titer in immunized and control group mice splenic cell culture supernatant after incubation in RPMI media. (b) ELISA reaction of the pooled sera and splenic lymphocyte culture supernatant of immunized group against eluted antigens of different molecular weights separated by SDS-PAGE shown with highest absorbance recorded against protein with MW 30-40 kDa.
Information

