Recent advancements in the study of human behavior, along with significant technological progress, have provided a more complete understanding of the neural circuits involved in aggressive responses to external stimuli. Notably, aggression as a voluntary behavior differs from reactive aggression in its neural connections and cerebral connectomics. Aggression is identified as part of a series of responses to stimuli that pose a potential threat to an individual’s physical integrity. By identifying the neural pathways involved in pathological aggression, we can modify this behavior by disrupting these pathways through functional neurosurgery. This study aims to demonstrate, through neuronal connectomics, the effects of postsurgical disconnection following functional neurosurgical procedures designed to treat aggression disorders. For many years, brain function was believed to result solely from the activity of specific cortical areas, which conditioned cognitive responses. However, recent advances in neurology, neuroimaging, neuropsychology, and neurosurgery have shifted our understanding of brain function, revealing a more complex network of connectivity. Despite we are now able to predict the location of primary cortical areas, patients may still experience unanticipated deficits in functions like judgment or memory after surgery. This suggests that traditionally silent regions of the brain may be more anatomically intricate and functionally redundant than previously understood. One of the significant advancements in neuroimaging is Diffusion Tensor Imaging (DTI), which has revolutionized psychiatric surgery, neuroendoscopy, and neuro-oncology. DTI enabled the formation of the Human Connectome Project (HCP), a large-scale initiative that provides detailed data on the brain's connectivity in healthy individuals. The most recent HCP findings have reclassified cortical regions previously described by Brodmann, based on functional connectivity, myelination, and cortical thickness. These advancements contribute to a much more detailed framework for studying brain function and its anatomical organization. This paper explores how these developments in connectomics and neuroimaging are being applied to the understanding and treatment of aggression disorders through functional neurosurgery.
| Published in | International Journal of Neurosurgery (Volume 9, Issue 1) |
| DOI | 10.11648/j.ijn.20250901.11 |
| Page(s) | 1-9 |
| Creative Commons |
This is an Open Access article, distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution and reproduction in any medium or format, provided the original work is properly cited. |
| Copyright |
Copyright © The Author(s), 2025. Published by Science Publishing Group |
Aggression Disorder, Functional Neurosurgery, Human Connectome Project (HCP), Neurosurgery and Aggression, Aggressive Behavior Treatment, Brain Connectomics
MRI | Magnetic Resonance Imaging |
DTI | Diffusion Tensor Imaging |
HCP | Human Connectome Project |
rfMRI | Resting-state Functional MRI |
tfMRI | Task-evoked Functional MRI |
dMRI | Diffusion Imaging MRI |
ET | Entorrinal |
TO | Temporoccipital |
TA | Anterior Temporal |
ACC | Anterior Cingular Cortex |
MCC | Middle Cingular Cortex |
PCC | Posterior Cingular Cortex |
RSC | Retrosplenial Cingular Cortex |
CB | Cingulum Bundle |
SPL | Superior Parietal Lobe |
SFG | Superior Frontal Gyrus |
SLF | Superior Longitudinal FasciculusHuman |
Para-CB | Paracingulate Portion |
Supra-CB | Supracingulate Portion |
| [1] | Baker CM, Burks JD, Briggs RG, Conner AK, Glenn CA, Sali G, McCoy TM, Battiste JD, O'Donoghue DL, Sughrue ME. A Connectomic Atlas of the Human Cerebrum-Chapter 1: Introduction, Methods, and Significance. Oper Neurosurg (Hagerstown). 2018 Dec 1; 15(suppl_1): S1-S9. PMID: 30260422; PMCID: PMC6887907. |
| [2] | Versace A, Graur S, Greenberg T, et al. Reduced focal fiber collinearity in the cingulum bundle in adults with obsessive-compulsive disorder. Neuropsychopharmacology. 2019; 44(7): 1182-1188. |
| [3] | Garcia-Garcia S, Kakaizada S, Oleaga L, Benet A, Rincon-Toroella J, González-Sánchez JJ. Presurgical simulation for neuroendoscopic procedures: virtual study of the integrity of neurological pathways using diffusion tensor imaging tractography. Neurol India. 2019; 67(3): 763-769. |
| [4] | Ozgural O, Al-Beyati ESM, Kahilogullari G. MR navigation and tractography-assisted transcranial neuroendoscopic aspiration of pediatric thalamic abscess. Pediatr Neurosurg. 2019; 54(5): 354-358. |
| [5] | Wu JS, Zhou LF, Tang WJ, et al. Clinical evaluation and follow-up outcome of diffusion tensor imaging-based functional neuronavigation: a prospective, controlled study in patients with gliomas involving pyramidal tracts. Neurosurgery. 2007; 61(5): 935-948; discussion 948-9. |
| [6] | Marcus D, Harwell J, Olsen T, et al. Informatics and data mining tools and strategies for the human connectome project. Front Neuroinform. 2011; 5: 4. |
| [7] | Glasser MF, Coalson TS, Robinson EC, et al. A multi-modal parcellation of human cerebral cortex. Nature. 2016; 536(7615): 171-178. |
| [8] | Amunts K, Zilles K. Architectonic Mapping of the Human Brain beyond Brodmann. Neuron. 2015 Dec 16; 88(6): 1086-1107. PMID: 26687219. |
| [9] | Brodmann K. Vergleichende Lokalisationslehre der Grosshirnrinde in ihren Prinzipien dargestellt auf Grund des Zellenbaues. Barth; 1909. |
| [10] | Vogt C, Vogt O. Allgemeine ergebnisse unserer hirnforschung. JA Barth; 1919. |
| [11] | von Economo CF, Koskinas GN. Die cytoarchitektonik der hirnrinde des erwachsenen menschen. J. Springer; 1925. |
| [12] | Baker CM, Burks JD, Briggs RG, Conner AK, Glenn CA, Robbins JM, Sheets JR, Sali G, McCoy TM, Battiste JD, O'Donoghue DL, Sughrue ME. A Connectomic Atlas of the Human Cerebrum-Chapter 5: The Insula and Opercular Cortex. Oper Neurosurg (Hagerstown). 2018 Dec 1; 15(suppl_1): S175-S244. PMID: 30260456; PMCID: PMC6924540. |
| [13] | Jennings, Jonathan E MD; Kassam, Amin B MD; Fukui, Melanie B MD; Monroy-Sosa, Alejandro MD; Chakravarthi, Srikant MD; Kojis, Nathan MBA; Rovin, Richard A MD. The Surgical White Matter Chassis: A Practical 3-Dimensional Atlas for Planning Subcortical Surgical Trajectories. Operative Neurosurgery 14(5): p 469-482, May 2018. | |
| [14] | Sporns O, Chialvo DR, Kaiser M, Hilgetag CC. Organization, development and function of complex brain networks. Trends Cogn Sci. 2004; 8(9): 418-425. |
| [15] | Bassett DS, Gazzaniga MS. Understanding complexity in the human brain. Trends Cogn Sci. 2011; 15(5): 200-209. |
| [16] | Hart, Michael G., Rolf J. F. Ypma, Rafael Romero-Garcia, Stephen J. Price, and John Suckling. "Graph theory analysis of complex brain networks: new concepts in brain mapping applied to neurosurgery". Journal of Neurosurgery JNS 124.6 (2016): 1665-1678. |
| [17] | Martin R. Understanding Memory Systems in TLE: Networks, Nodes, and Hubs. Epilepsy Curr. 2016 May-Jun; 16(3): 153-5. PMID: 27330439; PMCID: PMC4913845. |
| [18] | Elam JS, Glasser MF, Harms MP, Sotiropoulos SN, Andersson JLR, Burgess GC, Curtiss SW, Oostenveld R, Larson-Prior LJ, Schoffelen JM, Hodge MR, Cler EA, Marcus DM, Barch DM, Yacoub E, Smith SM, Ugurbil K, Van Essen DC. The Human Connectome Project: A retrospective. Neuroimage. 2021 Dec 1; 244: 118543. Epub 2021 Sep 8. PMID: 34508893; PMCID: PMC9387634. |
| [19] | Wu Y, Sun D, Wang Y, Wang Y, ou S. Segmentation of the Cingulum Bundle in the Human Brain: A New Perspective Based on DSI Tractography and Fiber Dissection Study. Front Neuroanat. 2016 Sep 7; 10: 84. PMID: 27656132; PMCID: PMC5013069. |
| [20] | Agrawal, A., Kapfhammer, J. P., Kress, A., Wichers, H., Deep, A., Feindel, W., et al. (2011). Josef Klingler's models of white matter tracts: influences on neuroanatomy, neurosurgery and neuroimaging. Neurosurgery 69, 238-252; discussion 252-234. |
| [21] | Vogt, B. A. (2016). Midcingulatecortex: structure, connections, homologies, functions anddiseases. J. Chem. Neuroanat. 74, 28-46. |
| [22] | Vogt, B. A., Derbyshire, S., and Jones, A. K. (1996). Pain processing in four regions ofhuman cingulate cortex localized with co-registered PET and MR imaging. Eur. J. Neurosci. 8, 1461-1473. |
| [23] | Vogt, B. A., Finch, D. M., and Olson, C. R. (1992). Functional heterogeneity in cingulate cortex: the anterior executive and posterior evaluative regions. Cereb. Cortex 2, 435-443. |
| [24] | Vogt, B. A., and Laureys, S. (2005). Posterior cingulate, precunealand retrosplenial cortices: cytology and components of the neural network correlates ofconsciousness. Prog. BrainRes. 150, 205-217. |
| [25] | Bystritsky A, Spivak NM, Dang BH, Becerra SA, Distler MG, Jordan SE, Kuhn TP. Brain circuitry underlying the ABC model of anxiety. J Psychiatr Res. 2021 Jun; 138: 3-14. Epub 2021 Mar 25. PMID: 33798786. |
| [26] | Hernández Salazar M, Zarate Méndez A, Meneses Luna O, Ledesma Torres L, Paniagua Sierra R, Sánchez Moreno MC, Serrato Avila JL. Ablative stereotactic neurosurgery for irreducible neuroaggressive disorder in pediatric patients. Neurosurgery (Astur: Engl Ed). 2018 Nov-Dec; 29(6): 296-303. English, Spanish. Epub 2018 Jun 18. PMID: 29914842. |
| [27] | Bogerts B, Schöne M, Breitschuh S. Brain alterations potentially associated with aggression and terrorism. CNS Spectr. 2018 Apr; 23(2): 129-140. Epub 2017 Aug 14. PMID: 28803592. |
APA Style
Donath, J. L. C., Delgado, J. M. R., Luna, O. M., Salazar, M. H. (2025). Impact of Postsurgical Disconnection in Aggression Disorder Treatment, Latin American Single Center Experience: A Neuronal Connectomics Analysis. International Journal of Neurosurgery, 9(1), 1-9. https://doi.org/10.11648/j.ijn.20250901.11
ACS Style
Donath, J. L. C.; Delgado, J. M. R.; Luna, O. M.; Salazar, M. H. Impact of Postsurgical Disconnection in Aggression Disorder Treatment, Latin American Single Center Experience: A Neuronal Connectomics Analysis. Int. J. Neurosurg. 2025, 9(1), 1-9. doi: 10.11648/j.ijn.20250901.11
@article{10.11648/j.ijn.20250901.11,
author = {José Luis Capellini Donath and José Miguel Ramos Delgado and Oscar Meneses Luna and Manuel Hernández Salazar},
title = {Impact of Postsurgical Disconnection in Aggression Disorder Treatment, Latin American Single Center Experience: A Neuronal Connectomics Analysis
},
journal = {International Journal of Neurosurgery},
volume = {9},
number = {1},
pages = {1-9},
doi = {10.11648/j.ijn.20250901.11},
url = {https://doi.org/10.11648/j.ijn.20250901.11},
eprint = {https://article.sciencepublishinggroup.com/pdf/10.11648.j.ijn.20250901.11},
abstract = {Recent advancements in the study of human behavior, along with significant technological progress, have provided a more complete understanding of the neural circuits involved in aggressive responses to external stimuli. Notably, aggression as a voluntary behavior differs from reactive aggression in its neural connections and cerebral connectomics. Aggression is identified as part of a series of responses to stimuli that pose a potential threat to an individual’s physical integrity. By identifying the neural pathways involved in pathological aggression, we can modify this behavior by disrupting these pathways through functional neurosurgery. This study aims to demonstrate, through neuronal connectomics, the effects of postsurgical disconnection following functional neurosurgical procedures designed to treat aggression disorders. For many years, brain function was believed to result solely from the activity of specific cortical areas, which conditioned cognitive responses. However, recent advances in neurology, neuroimaging, neuropsychology, and neurosurgery have shifted our understanding of brain function, revealing a more complex network of connectivity. Despite we are now able to predict the location of primary cortical areas, patients may still experience unanticipated deficits in functions like judgment or memory after surgery. This suggests that traditionally silent regions of the brain may be more anatomically intricate and functionally redundant than previously understood. One of the significant advancements in neuroimaging is Diffusion Tensor Imaging (DTI), which has revolutionized psychiatric surgery, neuroendoscopy, and neuro-oncology. DTI enabled the formation of the Human Connectome Project (HCP), a large-scale initiative that provides detailed data on the brain's connectivity in healthy individuals. The most recent HCP findings have reclassified cortical regions previously described by Brodmann, based on functional connectivity, myelination, and cortical thickness. These advancements contribute to a much more detailed framework for studying brain function and its anatomical organization. This paper explores how these developments in connectomics and neuroimaging are being applied to the understanding and treatment of aggression disorders through functional neurosurgery.
},
year = {2025}
}
TY - JOUR T1 - Impact of Postsurgical Disconnection in Aggression Disorder Treatment, Latin American Single Center Experience: A Neuronal Connectomics Analysis AU - José Luis Capellini Donath AU - José Miguel Ramos Delgado AU - Oscar Meneses Luna AU - Manuel Hernández Salazar Y1 - 2025/02/20 PY - 2025 N1 - https://doi.org/10.11648/j.ijn.20250901.11 DO - 10.11648/j.ijn.20250901.11 T2 - International Journal of Neurosurgery JF - International Journal of Neurosurgery JO - International Journal of Neurosurgery SP - 1 EP - 9 PB - Science Publishing Group SN - 2640-1959 UR - https://doi.org/10.11648/j.ijn.20250901.11 AB - Recent advancements in the study of human behavior, along with significant technological progress, have provided a more complete understanding of the neural circuits involved in aggressive responses to external stimuli. Notably, aggression as a voluntary behavior differs from reactive aggression in its neural connections and cerebral connectomics. Aggression is identified as part of a series of responses to stimuli that pose a potential threat to an individual’s physical integrity. By identifying the neural pathways involved in pathological aggression, we can modify this behavior by disrupting these pathways through functional neurosurgery. This study aims to demonstrate, through neuronal connectomics, the effects of postsurgical disconnection following functional neurosurgical procedures designed to treat aggression disorders. For many years, brain function was believed to result solely from the activity of specific cortical areas, which conditioned cognitive responses. However, recent advances in neurology, neuroimaging, neuropsychology, and neurosurgery have shifted our understanding of brain function, revealing a more complex network of connectivity. Despite we are now able to predict the location of primary cortical areas, patients may still experience unanticipated deficits in functions like judgment or memory after surgery. This suggests that traditionally silent regions of the brain may be more anatomically intricate and functionally redundant than previously understood. One of the significant advancements in neuroimaging is Diffusion Tensor Imaging (DTI), which has revolutionized psychiatric surgery, neuroendoscopy, and neuro-oncology. DTI enabled the formation of the Human Connectome Project (HCP), a large-scale initiative that provides detailed data on the brain's connectivity in healthy individuals. The most recent HCP findings have reclassified cortical regions previously described by Brodmann, based on functional connectivity, myelination, and cortical thickness. These advancements contribute to a much more detailed framework for studying brain function and its anatomical organization. This paper explores how these developments in connectomics and neuroimaging are being applied to the understanding and treatment of aggression disorders through functional neurosurgery. VL - 9 IS - 1 ER -
Department of Neurosurgery, Centro Médico Nacional “20 de Noviembre” ISSSTE, Mexico City, Mexico
Department of Neurosurgery, Centro Médico Nacional “20 de Noviembre” ISSSTE, Mexico City, Mexico
Department of Psychiatry, Centro Médico Nacional “20 de Noviembre” ISSSTE, Mexico City, Mexico
Department of Neurosurgery, Centro Médico Nacional “20 de Noviembre” ISSSTE, Mexico City, Mexico
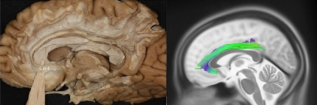
Figure 1. Cingulate fasciculus CBI segment, tractography, and fiber dissection.
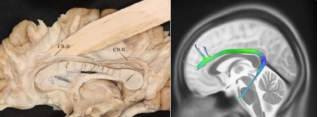
Figure 2. CBII path calculated by tractography and demonstrated with fiber dissection.
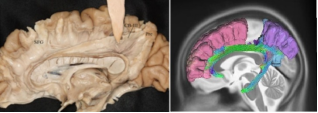
Figure 3. CBIII and its trajectory by tractography and fiber dissection.
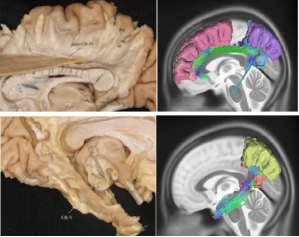
Figure 4. CB IV and CB V segment demonstrated by fiber dissection and tractography.
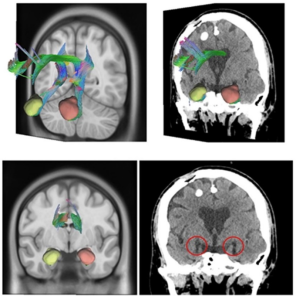
Figure 5. Fusion of postoperative tomography with tractography extracted from HCP demonstrating the interruption of tonsillar fibers in ablation of the bilateral tonsillar complex. Top left image: 3D reconstruction of the isolated bilateral amygdala complex and complete bilateral cingulate tract in DSI Studio Software in coronal plane. Top right image: Coronal section of a patient with bilateral ablation of the tonsillar complex, medial portion, postoperative image. Bottom left image: 3D reconstruction of the isolated bilateral tonsillar complex and complete bilateral cingulate tract in DSI Studio Software in an oblique coronal plane 3/4 to the right. Lower right image: Fusion of reconstruction of bilateral tonsillar complex and cingulate tract and postoperative image of a patient with bilateral tonsillar ablation and 3/4 oblique coronal plane cingulotomy rotated to the right where the interruption of the bilateral and cingulate medial tonsillar fibers is demonstrated.
Information

