Chemical insecticides have greatly contributed to the emergence of resistant mosquito populations in sub-Saharan Africa. Innovative strategies exploring novel animal-derived secretions may offer new avenues for effective vector control. We investigated the chemical composition of the hydroethanolic extract (HE) from millipede Ophistreptoides sp. and assessed its repellent and insecticidal effects against the susceptible Anopheles gambiae Kisumu strain. Millipedes were collected from vegetation and macerated in 70% distilled alcohol and 30% tap water for 5 days. A spectrophotometric approach was performed for the determination of all compounds present in the extract. The repellent effect was evaluated on fifty adult females of An. gambiae using impregnated papers with extract at different concentrations of 25, 50, 75, and 100 mg/mL, with acetone as control. The adulticidal effect was measured after 1 hour of exposure, with mortality assessed at 24, 48, and 72 hours post-exposure. Spectrophotometric analysis identified 2-2-methoxy-3-methyl-1,4-benzoquinone and methyl-1,4-benzoquinone as the main compounds in the extract. The repellency rate ranged from 57.69 ± 11. 46 at 25 mg/mL concentration to 97.54±1.42% at 100 mg/mL concentration. Mosquito mortality rates increased significantly (p < 0.05) with both exposure time and tested concentration with the highest toxicity recorded at 72 h post-exposure (LC50 = 18.09 mg/mL; LC95 = 186.44 mg/mL). This study highlights the potential of millipede-derived bioactive compounds in integrated vector management and suggests further research into their molecular modes of action and formulation for mosquito nets and spray.
| Published in | Research and Innovation (Volume 2, Issue 3) |
| DOI | 10.11648/j.ri.20260203.11 |
| Page(s) | 214-223 |
| Creative Commons |
This is an Open Access article, distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution and reproduction in any medium or format, provided the original work is properly cited. |
| Copyright |
Copyright © The Author(s), 2026. Published by Science Publishing Group |
Anopheles gambiae, Biological Control, Animal-derived Secretions, Benzoquinones, Vector Resistance, Bio-insecticide
Concentrations (mg/mL) | Times (minutes) | F | p | ||||||
|---|---|---|---|---|---|---|---|---|---|
<10 | 10 | 20 | 30 | 40 | 50 | 60 | |||
0 | 0,0 ± 0,0bA | 0,0 ± 0,0 d A | 0,0 ± 0,0 c A | 0,0 ± 0,0 c A | 0,0 ± 0,0 c A | 0,0 ± 0,0 c A | 0,0 ± 0,0 c A | - | - |
25 | 100± 0,0 a | 78,48 ± 3,54 c A | 68,00 ± 8,28 b A | 71,38 ± 3,55 b A | 75,64 ± 5,18 b A | 65,00 ± 8,66 b A | 57,69 ± 11,46b A | 1,04 | 0,42 |
50 | 100± 0,0aB | 85,45 ± 3,14 bc A | 86,79 ± 0,0 ab A | 85,62 ± 1,88 a A | 85,16 ± 0,31 ab A | 82,31 ± 1,33 ab A | 82,17 ± 2,66 ab A | 0,96 | 0,46 |
75 | 100± 0,0aB | 93,89 ± 2,55 ab A | 92,31 ± 4,43 a A | 88,27 ± 4,77 a A | 91,46 ± 3,95 a A | 87,37 ± 4,25 a A | 87,32 ± 4,22 a A | 0,47 | 0,79 |
100 | 100± 0,0aB | 97,54 ± 1,42 a A | 96,43 ± 4,12 a A | 94,61 ± 2,13 a A | 96,43 ± 2,06 a A | 94,61 ± 2,13 a A | 93,70 ± 1,61 a A | 0,57 | 0,71 |
F (4, 15) | - | 263,34 | 86,08 | 173,79 | 168,76 | 74,32 | 46,32 | ||
p | - | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | <0,001 | ||
Concentrations (mg/mL) | Mortality rate (%) | ||||
|---|---|---|---|---|---|
24h | 48h | 72h | p | F | |
25 | 29 ± 6,574 a B | 44 ± 5,416 c AB | 62 ± 3,887 c A | 0,001 | 9,342 |
50 | 42 ± 4,422 a B | 50 ± 2,981 bc B | 73 ± 4,485 bc A | <0,0001 | 16,002 |
75 | 47 ± 4,955 a B | 64 ± 5,416 ab A | 79 ± 3,480 b A | <0,0001 | 11,652 |
100 | 48 ± 4,422 a C | 76 ± 1,633 a B | 94 ± 1,633 a A | <0,0001 | 64,768 |
P | 0,051 | <0,0001 | <0,0001 | ||
F | 2,857 | 11,193 | 14,240 | ||
Observation time | R² | Slope ± ES | LC50 (IC) | LC95 (IC) | X² |
|---|---|---|---|---|---|
24 h | 0,161 | 0,859 ± 0,090 | 99,632 (67,729-462,620) | 8175,247 (1018,119-1,552E9) | 422,095*** |
48 h | 0,484 | 1,352 ± 0,090 | 37,800 (27,170-46,487) | 622,639 (310,301-2884,832) | 304,348*** |
72 h | 0,529 | 1,624 ± 0,099 | 18,094 (9,783-24,770) | 186,443 (126,800-405,935) | 321,065*** |
R2 | Coefficient of Determination |
CI | Confidence Interval |
CL | Lethal Dose |
ES | Standard Erro |
χ² | Chi-square |
| [1] |
World Health Organization (2025) Malaria end with us: reinvest. reimagine. reignite.
https://www/who.int/campaigns/world-malaria-day/2025 (Accessed on Sunday. 27 April. 2025). |
| [2] | Tankoua-Tchounda R. 2025 Gastrointestinal and blood-borne parasitic coinfections and HIV: effects on the immune system in patients admitted to the Bafoussam Regional Hospital (West Cameroon). Ph.D. Doctoral Thesis, Department of Animal Organism Biology, University of Douala, Cameroon: 160 p. |
| [3] | Fondjo E., Toto, JC., Tchouakui M., Eyisap WE, Patchoke S, Menze B, Chabi, J. High vector diversity and malaria transmission dynamics in five sentinel sites in Cameroon. Malar J. 2023; 22(1), 123. |
| [4] | Antonio-Nkondjio C, Ndo, C, Njiokou F., Bigoga JD, Awono-Ambene P, Etang J, Wondji, CS. Review of malaria situation in Cameroon: technical viewpoint on challenges and prospects for disease elimination. Parasit Vectors. 2019; 12(1), 501. |
| [5] | Mbakop LR., Awono-Ambene PH, Mandeng SE, Ekoko WE, Fesuh BN, Antonio-Nkondjio C., Etang, J. Malaria transmission around the Memve’ele hydroelectric dam in South Cameroon: a combined retrospective and prospective study, 2000–2016. IJERPH. 2019; 16(9), 1618. |
| [6] | Chouakeu NAK, Tchuinkam T, Bamou R., Bindamu MM, Talipouo A, Kopya, E, et al. Malaria transmission pattern across the Sahelian, humid savanna, highland and forest eco-epidemiological settings in Cameroon. Malar J. 2023; 22(1): 116. |
| [7] | Ekoko EW, Awono-Ambene P, Bigoga J, et al. Pattersn of anapheline feeding: resting behaviour and Plasmadium infections in North Cameroon, 2011-2014: Implications for malaria control. Parasit Vectors. 2019; 12(1), 297. |
| [8] | Simard F, Ayala D, Kamdem GC, Pombi M, Etouna J, Ose, K, et al. Ecological niche partitioning between Anopheles gambiae molecular forms in Cameroon: the ecological side of speciation. BMC Ecol. 2009; 9(1), 17. |
| [9] | World Health Organization (2023). Guidelines for laboratory and field-testing of long-lasting insecticidal nets. Geneva. (Accessed on Sunday. 27 April. 2025). |
| [10] | World Health Organization (WHO). (2021). “World Malaria Report 2021.” (Accessed on Sunday. 27 March 2024). |
| [11] | Antonio-Nkondjio C, Fossog BT, Ndo C, Djantio BM, Togouet SZ, Awono-Ambene P, al. Anopheles gambiae distribution and insecticide resistance in the cities of Douala and Yaoundé (Cameroon): influence of urban agriculture and pollution. Malar J. 2011; 10(1), 154. |
| [12] | Bamou, R, Sonhafouo-Chiana N, Mavridis K, Tchuinkam T, Wondji CS, Vontas J, et al. Status of insecticide resistance and its mechanisms in Anopheles gambiae and Anopheles coluzzii populations from forest settings in south Cameroon. Genes. 2019; 10(10): 741. |
| [13] | Nwane P, Etang J, Chouaїbou, M. et al. Multiple insecticide resistance mechanisms in Anopheles gambiae s.l. populations from Cameroon, Central Africa. Parasit Vectors. 2013; 6, 41. |
| [14] | Talipouo A, Mavridis K, Nchoutpouen E, et al. High insecticide resistance mediated by different mechanisms in Culex quinquefasciatus populations from the city of Yaoundé, Cameroon. Sci Rep. 2021; 11, 7322. |
| [15] | Sonhafouo-Chiana N, Nkahe LD, Kopya E, et al. Rapid evolution of insecticide resistance and patterns of pesticides usage in agriculture in the city of Yaoundé, Cameroon. Parasit Vectors. 2022; 15, 186. |
| [16] | Efa S, Elanga-Ndille E, Poumachu Y, Tene B, Mikande JZ, Zakaraiarou, Wondji CS, et al. Insecticide Resistance Profile and Mechanisms in An. gambiae s.l. from Ebolowa, South Cameroon. Insects. 2022, 13(12), 1133; |
| [17] | Thiomela RF, Tchouakui M, Menze BD, Nchoutpouen E, Ngongang-Yipmo ES, et al. Indoor residual spraying of experimental huts in Cameroon higlights the potential of Fludora® Fusion to control wild pyrethroid-resistant malaria vectors. BMC Infect Dis. 2024; 24(1): 733. |
| [18] | Smith J, Brown A, Green T. Neuroprotective properties of millipede secretions: Implications for antiepileptic drug development. J Chem Ecol. 2021; 47(3). 345-356. |
| [19] | Mbenoun Masse PS. An annotated checklist of the millipedes (Myriapoda Diplopoda) of Cameroon. with several new species and distribution records. Biodivers J. 2023; 14, 275-291. |
| [20] | Hopkin SP. Biology of the springtails (Insecta: Collembola) and their role in the soil ecosystem. Springer. 2015. |
| [21] | Mbenoun Masse PS. Makon SD. Tenyam Nkoh PB, Mwabvu T. Geographic distribution. diversity and conservation status of giant millipedes in southern Cameroon rainforest. Afr J Ecol. 2022; 60(4), 1071–1080. |
| [22] | Shear WA. The chemical defenses of millipedes (diplopoda): Biochemistry. physiology and ecology. Biochem Syst Ecol. 2015; 61, 78−117. |
| [23] | Bodner M. Vagalinski B, Makarov SE, Antić DŽ, Vujisić LV, Leis HJ, Raspotnig G. “Quinone Millipedes” Reconsidered: Evidence for a MosaicLike Taxonomic Distribution of Phenol-Based Secretions across the Julidae. J Chem Ecol. 2016; 42, 249-258. |
| [24] | Wood W, Hanke F, Kubo I, Carroll J, Crews P. Buzonamine. a new alkaloid from the defensive secretion of the millipede Buzonium crassipes. Biochem Syst Ecol. 2000; 28(4), 305-312. |
| [25] | Kandeda KA, Mbenoun MPS, Ateufack LB, Penda MZ, Baldagai N. Therapeutic potential of the extract of giant millipede Telodeinopus Canaliculatus on epileptogenesis and associated anxiety-like behaviour. Metab Brain Dis. 2024; 40-65. |
| [26] | Peckre LR, Defolie C, Kappeler PM, Fichtel C. Potential self -medication using millipede secretions in red-fronted lemur: combining anointment and ingestion for joint action against gastrointestinal parasites. Primates. 2018; 59, 483-494. |
| [27] | Johnson R, Lee M. Chemical ecology of millipede secretions: Efficacy as insect repellents. Toxins. 2022; 14(5), 321-334. |
| [28] | Garcia L, Patel S, Kim H. Lethal effects of millipede toxins on insect predators: Ecological implications. J Insect Sci. 2023; 23(2), 112-124. |
| [29] | Herborne JB. Phytochemical methods. A Guide To Modern Techniques of Plant Analysis. 1973; 2, 5-11. |
| [30] | Sofowora EA. Phytochemical screening: Medicinal plant and traditional medicine in Africa. Spectrum Books Ltd. Ibadan. Nigeria; 1993; 270-289. |
| [31] | N’Guessan R, Corbel V, Akogbéto M, Rowland M. Reduced efficacy of insecticide-treated nets and indoor residual spraying for malaria control in pyrethroid resistance area. Benin. EID. 2007; 13(2), 199-206. |
| [32] | Corbel V, Fonseca DM, Weetman D, Pinto J, Achee NL, Chandre F, Coulibaly MB. et al. International workshop on insecticide resistance in vectors of arboviruses. December 2016. Rio de Janeiro. Brazil. Parasit Vectors. 2017; 10, 1-16. |
| [33] | Grieco JP, Achee NL, Sardelis MR, Chauhan KR, Roberts DR. A Novel high-throughput screening system to evaluate the behavioural response of adult mosquitoes to chemicals. J Am Mosq Control Assoc. 2005; 21(4), 404-411. |
| [34] | Abbott WS. A method of computing the effectiveness of an insecticide. J Econ Entomol. 1925; 18, 265-267. |
| [35] | Williams L, Singh P, Williams L. Biology and biological action of the defensive secretion from a Jamaican millipede. Nat Sci. 1997; 84(4), 143-144. |
| [36] | Buden D, Attygalle A, Wu X. Distribution of the Chuuk Islands giant millipede. Acladocricus setigerus (Spirobolida: Rhinocricidae). and identification of its defensive compounds. Pac Sci. 2004; 58(4), 625-636. |
| [37] | Makarov SE, Curcic BPM, Tesevic VV. Defensive secretions in three species of polydesmids (Diplopoda. Polydesmida. Polydesmidae). J Chem Ecol. 2010; 36, 978-982. |
| [38] | Eisner T, Meinwald J. Defensive secretions of arthropods. Science. 1966; 153, 1341-1350. |
| [39] | Monro A, Chadha M, Meinwald J, Eisner T. Defense mechanisms of arthropods. VI. Para-benzoquinones in the secretion of five species of millipedes. Anna Entomol Soc Am. 1962; 55, 261-262. |
| [40] | Deml R, Huth A. Benzoquinones and hydroquinones in defensive secretions of tropical millipedes. Die Naturwiss. 2000; 87(2), 80-82. |
| [41] | Wu X, Buden D, Attygalle A. Hydroquinones from defensive secretion of a giant Pacific millipede. V n (Diplopoda: Spirobolida). Chemoecology. 2007; 17, 131-138. |
| [42] | Kuwahara Y, Omura H, Tanabe, T. 2-Nitroethenylbezenes as natural products in millipede defense secretions. Die Naturwiss. 2002; 89(7), 308-310. |
| [43] | Kuwahara Y, Mori N, Tanabe T. Detection of a neotropical frog alkaloid spiropyrrolizidine 236 from a Japanese polyzoniid millipede Kiusiozonium okai as a major defense component together with polyzonimine and nitropolyzonamine. Japanese J Environ Entomol Zool. 2007; 18(2), 91-95. |
| [44] | White G. Terminology of Insect Repellents: Insect Repellents: Principles. Pp 31–46 in Debboun M, Frances S, Strickman D. (Eds). Methods and Uses. 2007, CRC Press. |
| [45] | Peschke K, EisnerT. Defensive secretion of the tenebrionid beetle. Blaps mucronata: physical and chemical determinants of effectiveness. J Comp Physiol. 1987; 161(3), 377-388. |
| [46] | Weldon P, Aldrich J, Klum OJ, Debboun M. Centipede benzoquinones deter mosquitoes and induce self-anointing in capuchin monkeys (Cebus spp). Naturwissenschaften. 2003; 90, 301-304. |
| [47] | Carroll JF, Kramer M, Weldon PJ, Robbins RG. Produits chimiques d'onction et ectoparasites: effets des benzoquinones des millepattes sur la tique étoilée solitaire [Chemical secretions and ectoparasites: Effects of millipede benzoquinones on the lone star tick]. Amblyomma americanum. J Chem Ecol. 2005; 31(1): 63-75. |
| [48] | Grünewald B, Siefert P. Acetylcholine and its receptors in the honeybee: involvement in development and impairment by neonicotinoids. Insects. 2019; 10(12), 420. |
| [49] | Naqvi SNH. Biological evaluation of fresh extracts and some neem components with reference to abnormalities and esterase activity in insects. In: Natural pesticides from the neem tree and other tropical plants. Proceeding of the 3rd international neem conference. Nairobi. Kenya. France. ANPP. 187; II (6): 315-330. |
| [50] | Pesewu GA, Cutler RR, Humber DP. Activité antibactérienne des plantes utilisées dans la médecine traditionnelle du Ghana avec une référence particulière au SARM [Antibacterial activity of plants used in traditional medicine in Ghana with a particular reference to MRSA]. J Ethnopharmacol. 2008; 116(1), 102-111. |
APA Style
Jordan, K. T. S., Philippe, N., Joseph, M. R., Didier, M. S., Serge, M. M. P. (2026). Potential Repellent and Insecticidal Effects of Millipede-derived Secretions Against Anopheles gambiae s.l., the Main Malaria Vector in Africa. Research and Innovation, 2(3), 214-223. https://doi.org/10.11648/j.ri.20260203.11
ACS Style
Jordan, K. T. S.; Philippe, N.; Joseph, M. R.; Didier, M. S.; Serge, M. M. P. Potential Repellent and Insecticidal Effects of Millipede-derived Secretions Against Anopheles gambiae s.l., the Main Malaria Vector in Africa. Res. Innovation 2026, 2(3), 214-223. doi: 10.11648/j.ri.20260203.11
@article{10.11648/j.ri.20260203.11,
author = {Kue Tagne Styve Jordan and Nwane Philippe and Mahob Raymond Joseph and Makon Samuel Didier and Mbenoun Massé Paul Serge},
title = {Potential Repellent and Insecticidal Effects of
Millipede-derived Secretions Against Anopheles gambiae s.l., the Main Malaria Vector in Africa},
journal = {Research and Innovation},
volume = {2},
number = {3},
pages = {214-223},
doi = {10.11648/j.ri.20260203.11},
url = {https://doi.org/10.11648/j.ri.20260203.11},
eprint = {https://article.sciencepublishinggroup.com/pdf/10.11648.j.ri.20260203.11},
abstract = {Chemical insecticides have greatly contributed to the emergence of resistant mosquito populations in sub-Saharan Africa. Innovative strategies exploring novel animal-derived secretions may offer new avenues for effective vector control. We investigated the chemical composition of the hydroethanolic extract (HE) from millipede Ophistreptoides sp. and assessed its repellent and insecticidal effects against the susceptible Anopheles gambiae Kisumu strain. Millipedes were collected from vegetation and macerated in 70% distilled alcohol and 30% tap water for 5 days. A spectrophotometric approach was performed for the determination of all compounds present in the extract. The repellent effect was evaluated on fifty adult females of An. gambiae using impregnated papers with extract at different concentrations of 25, 50, 75, and 100 mg/mL, with acetone as control. The adulticidal effect was measured after 1 hour of exposure, with mortality assessed at 24, 48, and 72 hours post-exposure. Spectrophotometric analysis identified 2-2-methoxy-3-methyl-1,4-benzoquinone and methyl-1,4-benzoquinone as the main compounds in the extract. The repellency rate ranged from 57.69 ± 11. 46 at 25 mg/mL concentration to 97.54±1.42% at 100 mg/mL concentration. Mosquito mortality rates increased significantly (p 50 = 18.09 mg/mL; LC95 = 186.44 mg/mL). This study highlights the potential of millipede-derived bioactive compounds in integrated vector management and suggests further research into their molecular modes of action and formulation for mosquito nets and spray.},
year = {2026}
}
TY - JOUR T1 - Potential Repellent and Insecticidal Effects of Millipede-derived Secretions Against Anopheles gambiae s.l., the Main Malaria Vector in Africa AU - Kue Tagne Styve Jordan AU - Nwane Philippe AU - Mahob Raymond Joseph AU - Makon Samuel Didier AU - Mbenoun Massé Paul Serge Y1 - 2026/05/08 PY - 2026 N1 - https://doi.org/10.11648/j.ri.20260203.11 DO - 10.11648/j.ri.20260203.11 T2 - Research and Innovation JF - Research and Innovation JO - Research and Innovation SP - 214 EP - 223 PB - Science Publishing Group SN - 3070-6297 UR - https://doi.org/10.11648/j.ri.20260203.11 AB - Chemical insecticides have greatly contributed to the emergence of resistant mosquito populations in sub-Saharan Africa. Innovative strategies exploring novel animal-derived secretions may offer new avenues for effective vector control. We investigated the chemical composition of the hydroethanolic extract (HE) from millipede Ophistreptoides sp. and assessed its repellent and insecticidal effects against the susceptible Anopheles gambiae Kisumu strain. Millipedes were collected from vegetation and macerated in 70% distilled alcohol and 30% tap water for 5 days. A spectrophotometric approach was performed for the determination of all compounds present in the extract. The repellent effect was evaluated on fifty adult females of An. gambiae using impregnated papers with extract at different concentrations of 25, 50, 75, and 100 mg/mL, with acetone as control. The adulticidal effect was measured after 1 hour of exposure, with mortality assessed at 24, 48, and 72 hours post-exposure. Spectrophotometric analysis identified 2-2-methoxy-3-methyl-1,4-benzoquinone and methyl-1,4-benzoquinone as the main compounds in the extract. The repellency rate ranged from 57.69 ± 11. 46 at 25 mg/mL concentration to 97.54±1.42% at 100 mg/mL concentration. Mosquito mortality rates increased significantly (p 50 = 18.09 mg/mL; LC95 = 186.44 mg/mL). This study highlights the potential of millipede-derived bioactive compounds in integrated vector management and suggests further research into their molecular modes of action and formulation for mosquito nets and spray. VL - 2 IS - 3 ER -
Department of Animal Biology and Physiology, University of Yaounde 1, Yaounde, Cameroon
Department of Animal Biology and Physiology, University of Yaounde 1, Yaounde, Cameroon

Figure 1. Morphology of the spiraled Ophistreptoides sp. collected at the campus of the University of Yaounde I, Cameroon.
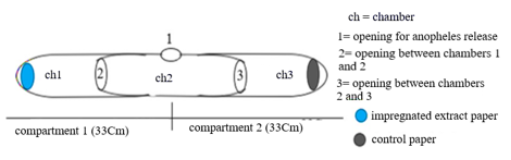
Figure 2. Schematic drawing of a modified repellency test of Grieco et al. (2005) including chambers and compartment legends.
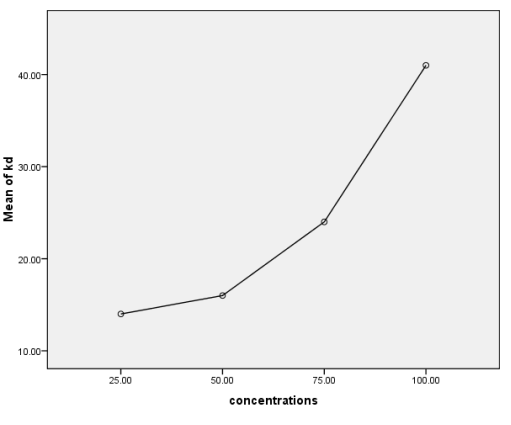
Figure 3. Percentage of mosquitoes exhibiting knockdown 1 hour after exposure.
Information

